Japanese
English
- 有料閲覧
- Abstract 文献概要
- 1ページ目 Look Inside
- 参考文献 Reference
[1] はじめに―アクチンによる形態形成と細胞運動
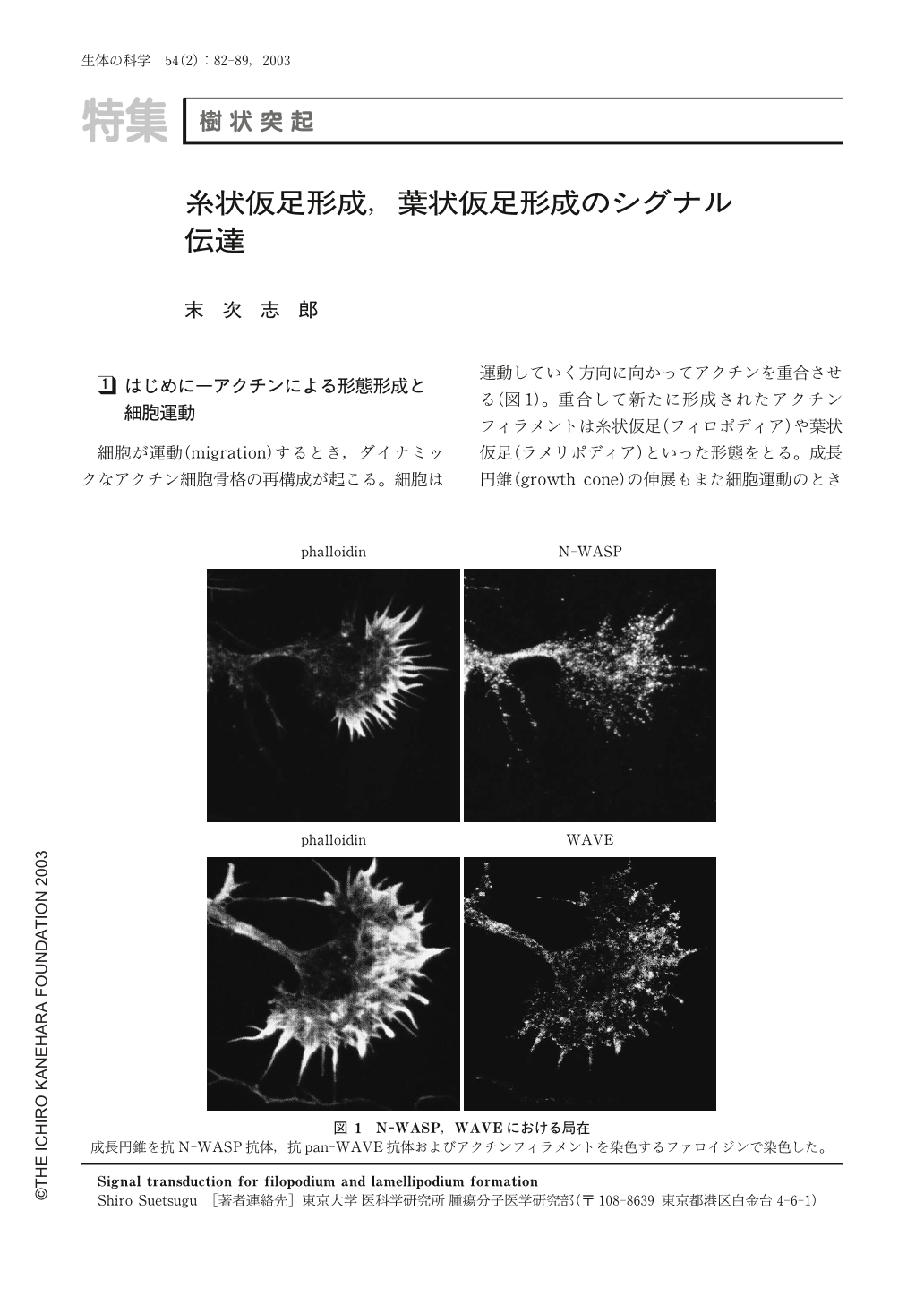
細胞が運動(migration)するとき,ダイナミックなアクチン細胞骨格の再構成が起こる。細胞は運動していく方向に向かってアクチンを重合させる(図1)。重合して新たに形成されたアクチンフィラメントは糸状仮足(フィロポディア)や葉状仮足(ラメリポディア)といった形態をとる。成長円錐(growth cone)の伸展もまた細胞運動のときと同様に糸状仮足や葉状仮足形成を伴う。糸状仮足はアクチンフィラメントの束からなる突起状の構造であり,葉状仮足は網目状のアクチンフィラメントからなっている。これらの構造の形成は非常にダイナミックであり,非常に早いアクチン重合が起こる必要がある。アクチンフィラメントは反矢じり端と矢じり端の二つの端を持つが,早く重合するのは反矢じり端である。このため,早いアクチン重合は反矢じり端で起こる。
アクチン重合を開始するためにはアクチン重合核を形成する必要がある。アクチンだけのin vitro系において,アクチン重合核形成はアクチンの三量体形成か,反矢じり端が露出されたアクチンフィラメントの付加である。しかしながら,細胞内では単量体アクチン結合タンパク質のためアクチンの三量体形成はほとんど起こらない。また,アクチンフィラメントの反矢じり端はキャッピングタンパク質など様々なアクチン結合タンパク質によって覆われている。そのため細胞におけるアクチン重合の開始はシグナル依存的なタンパク質が制御している。一つはArp2/3複合体という重合核になりうるタンパク質を活性化し,重合核とする方法である。もう一つは覆われている反矢じり端をPIP2などのシグナル分子が露出させ,この端を重合核として用いる方法である。
特に葉状仮足においては,Arp2/3複合体はアクチン重合核形成において重要であるだけでなく,網目状構造を作る基礎となる。in vitroでArp2/3複合体とアクチンを混ぜ重合条件にすると70度の角度で枝分かれしたアクチンフィラメントが形成される1)。これは細胞の葉状仮足でみられるアクチンの枝分かれ角度と同じである2)。また,葉状仮足においてArp2/3複合体は枝分かれの基部に存在している。アクチン重合を誘導するメカニズムはほかにも考えられるが,Arp2/3複合体を介したメカニズムが網目状の形態を直接説明できる。
このArp2/3複合体の活性化因子として重要なのがWASPファミリータンパク質である。WASPファミリータンパク質はArp2/3複合体のアクチン重合における重合核形成機能を活性化し,アクチン重合を爆発的に引き起こす3,4)。実際に葉状仮足形成においてWASPファミリータンパク質およびArp2/3複合体は必要不可欠であることが示されている5)。成長円錐にWASPファミリータンパク質は局在し,成長円錐の伸長におけるアクチン重合を制御していると考えられる(図1)。
仮足形成においてはこのような重合を促進するシグナルだけでなく,アクチン脱重合を抑制し,その結果としての重合の促進も重要であると考えられる。脱重合にはコフィリンと呼ばれるタンパク質が重要であると考えられ,コフィリンはLIMキナーゼによってシグナル依存的に制御されている。また近年,Ena/VASPファミリータンパク質が反矢じり端のキャッピングタンパク質と拮抗し,仮足形成におけるアクチン重合を促進しているとのデータが提出された。
本稿ではWASPファミリータンパク質とArp2/3複合体,コフィリンとLIMキナーゼ,Ena/VASPファミリータンパク質とキャッピングタンパク質について述べる。
Copyright © 2003, THE ICHIRO KANEHARA FOUNDATION. All rights reserved.

